Virus
I virus: parassiti obbligati delle cellule
I virus sono piccoli parassiti che necessitano di una cellula per poter completare il proprio ciclo vitale.
Per tale proprietà sono chiamati endoparassiti cellulari obbligati.
I virus sono in grado di infettare qualsiasi cellula: animale, vegetale e batterica.
I virus batterici sono detti batteriofagi o fagi.
Alcuni virus possono causare gravi patologie nell'uomo e negli animali, da qui il significato latino del termine virus, veleno.
I virus vegetali, trasmessi attraverso il morso degli insetti, possono provocare gravi danni ai raccolti.
In genere i virus vegetali non causano la morte della pianta, ma si limitano a rallentare la crescita, a indurre cambiamenti di colore, a provocare la comparsa di striature sui petali o alterazioni della superficie fogliare.
Ne è un esempio il virus del mosaico del tabacco (TMV), uno dei primi virus ad essere stato studiato e isolato, aprendo la strada alla scoperta e alla descrizione di queste particelle, la cui natura e origine evolutiva non sono state ancora del tutto chiarite.

Il virus del mosaico del tabacco (TMV) è stato il primo ad essere purificato nel 1935. Oltre al tabacco questo virus infetta pomodori e altre solanacee. In A e B si nota la tipica screziatura sulle foglie delle piante rispettivamente di tabacco e pomodoro. In C è possibile notare i cambiamenti di colore indotti anche sul frutto di pomodoro.
Organizzazione strutturale dei virus
I virus non hanno un'organizzazione cellulare, ma macromolecolare.
Non sono capaci di svolgere attività biosintetiche, essendo privi di ribosomi ad esempio, e metaboliche. Per tali ragioni essi si trovano al confine tra esseri viventi e non.
La definizione più esatta dei virus è quella di elementi genetici mobili in quanto trasportano, all'interno di un involucro proteico detto capside, una molecola di acido nucleico, DNA o RNA, che iniettata nella cellula ospite ne riprogramma e gestisce gli enzimi coinvolti nei processi trascrizionali e traduzionali, per produrre un gran numero di nuovi virus rilasciati poi all'esterno.
Dimensioni dei virus
Ogni virus presenta una forma extracellulare, metabolicamente inerte, detta virione, che è responsabile del riconoscimento della cellula ospite, nella quale inietta l'acido nucleico dando inizio alla fase intracellulare del suo ciclo vitale, che coincide con il processo infettivo vero e proprio.
I virus nella loro forma extracellulare presentano forme e dimensioni diverse.
Essi sono più piccoli delle cellule batteriche e le dimensioni variano tra 20 e 300 nanometri.
Solo la messa a punto del microscopio elettronico a partire dagli '40 del secolo scorso ne ha consentito l'osservazione.
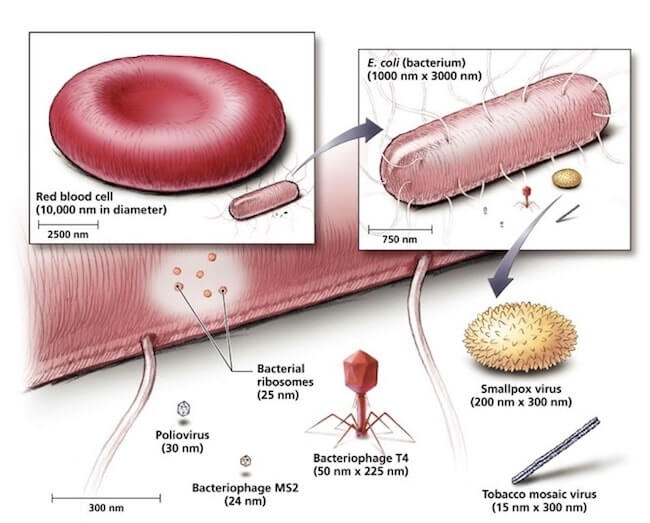
Confronto tra le dimensioni di una tipica cellula animale, il globulo rosso, il batterio Escherichia coli e alcuni virus, i più piccoli dei quali hanno dimensioni paragonabili a quelle dei ribosomi batterici.
Struttura dei virus
Il virione può essere descritto come l'insieme di acido nucleico e capside, cioè il nucleocapside.
Il capside si compone di più subunità proteiche, le quali, una volta prodotte nella cellula ospite, sono in grado di assemblarsi spontaneamente.
I virus dotati solo di nucleocapside sono detti nudi, molti sono invece rivestiti.
Nei virus rivestiti il nucleocapside è racchiuso da una membrana formata da un doppio strato lipidico, derivata dalla cellula ospite, in cui sono inserite glicoproteine (chiamate spicole) virus-specifiche di fondamentale importanza per il riconoscimento delle cellule ospiti e il rilascio delle nuove particelle virali. Questo ulteriore rivestimento è detto pericapside o envelope.
Sono dotati di envelope molti virus che infettano le cellule animali, alcuni delle piante e uno batterico.
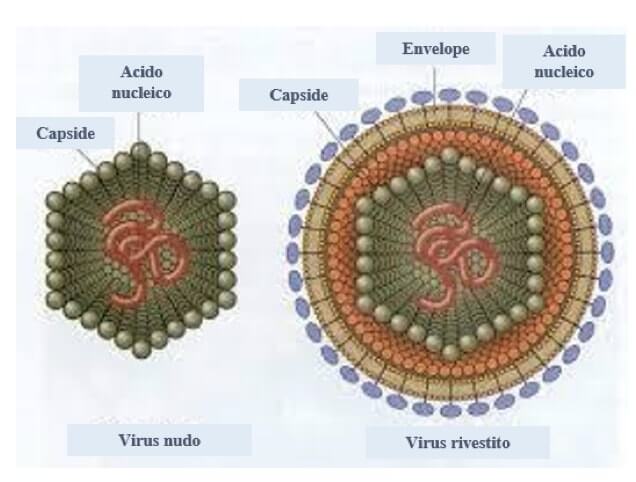
La struttura di base di tutti i virus prevede un rivestimento proteico, il capside, al cui interno si trova una molecola di acido nucleico, DNA o RNA. I virus dotati di solo nucleocapside sono detti nudi, quelli rivestiti presentano in aggiunta il pericapside o envelope, di natura lipidica e glicoproteica.
Simmetria dei virus
La disposizione delle subunità proteiche del capside dà luogo a simmetrie diverse, utili nell'identificazione e classificazione dei virus.
Le simmetrie principali sono due: elicoidale e icosaedrica.
I virus a simmetria elicoidale sono di forma bastoncellare, ad esempio il virus del mosaico del tabacco (TMV), mentre la simmetria icosaedrica conferisce una forma di tipo sferico, tipica degli adenovirus, responsabili di infezioni nelle vie respiratorie nell'uomo e negli animali.
Esistono simmetrie più complesse, tipiche di molti batteriofagi, in cui si distinguono più parti, come una testa e una coda, ognuna con una propria organizzazione strutturale.
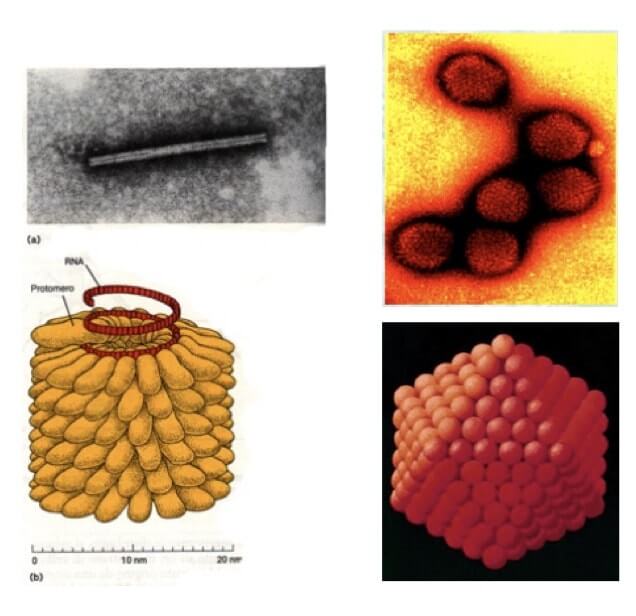
Le due principali simmetrie dei nucleocapsidi dei virus sono quella elicoidale (a sinistra) e quella icosaedrica (a destra). Le immagini in alto sono al microscopio elettronico.
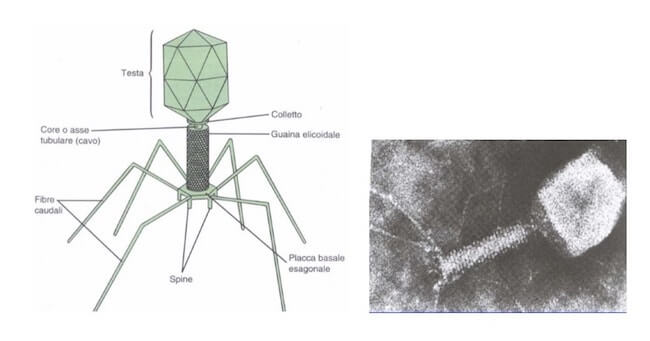
Le simmetrie più complesse si riscontrano nei batteriofagi, tipicamente formati da una testa icosaedrica e una coda elicoidale. Fibre caudali e spine sono coinvolte nel riconoscimento e nell'adesione sulla superficie delle cellule batteriche.
Classificazione dei virus
Nel tempo sono stati elaborati diversi criteri di classificazione dei virus.
Il sistema messo a punto dal medico Baltimore nel 1971 identifica sette classi in base alla tipologia di acido nucleico, DNA o RNA, alla configurazione a doppio o a singolo filamento e alle conseguenti strategie di replicazione.
Nel caso di genomi a RNA a singolo filamento, questo può essere a senso positivo o negativo a seconda se può fungere o meno da RNA messaggero (mRNA).
Alcuni virus utilizzano entrambe le tipologie di acido nucleico, ma in momenti diversi del ciclo replicativo (classi VI e VII).

Il sistema di classificazione dei virus messo a punto dal medico Baltimore nel 1971 prevede sette classi in funzione del tipo di acido nucleico e della configurazione a singolo o a doppio filamento. Il senso positivo o negativo del genoma a RNA si riferisce alla possibilità o meno di svolgere contemporaneamente la funzione di RNA messaggero.
Attualmente, l'organo ufficialmente incaricato della nomenclatura e classificazione dei virus è l'ITCV (Commissione Internazionale per la Classificazione dei Virus), che utilizza gli stessi criteri impiegati nella tassonomia degli organismi viventi, raggruppando i virus nelle categorie Ordine, Famiglia, Sottofamiglia, Genere e Specie.
Nella classificazione dei virus, oltre alla tipologia di DNA si prendono in considerazione anche il tipo di organismo infettato, la morfologia e la simmetria del capside, il meccanismo di replicazione.
Lo spettro d'ospite
La tipologia di organismi riconosciuti e infettati da un dato virus è detta spettro d'ospite.
In genere vi è un'elevata specificità tra virus e tipo di organismo infettato.
Nel caso di organismi pluricellulari (piante e animali) i virus riconoscono solo alcuni tipi di cellule o tessuti.
Questa affinità tessuto-specifica è detta tropismo e dipende dall'interazione tra molecole della superficie virale e di quella cellulare nella fase di riconoscimento della cellula ospite.
I poliovirus infettano e distruggono alcuni tipi di cellule nervose, i rinovirus (responsabili del comune raffreddore) sono in grado di moltiplicarsi nel tratto respiratorio superiore perché adattati alle condizioni di bassa temperatura ed elevata disponibilità di ossigeno, gli enterovirus prediligono le vie intestinali, mentre il virus HIV, responsabile dell'immunodeficienza acquisita (AIDS), agisce selettivamente sui linfociti T.
Sono note, tuttavia, alcune importanti eccezioni di virus in grado di infettare organismi anche non evolutivamente correlati.
I virus di alcune piante infettano anche gli insetti che si nutrono di esse, sfruttandoli come vettori da una pianta a un'altra.
Alcune gravi malattie infettive dell'uomo e di altri animali sono causate da virus trasmesse da insetti e capaci quindi di replicarsi sia in un ospite Vertebrato che in un Invertebrato.
La replicazione virale
Per infettare una cellula, un virus deve aderire alla sua superficie, penetrarvi all'interno e liberarsi degli involucri esterni per esporre il genoma agli enzimi della cellula, implicati nella sua replicazione e nell'espressione delle proteine codificate.
In ultimo, le nuove particelle virali assemblate nella cellula ospite devono essere liberate per poter infettare altre cellule.
1) Attacco
La fase di adesione alla cellula ospite è detta attacco o adsorbimento.
È affidata all'interazione tra molecole del capside o dell'envelope, definite nel complesso anti-recettori, ai recettori della cellula bersaglio.
Questi ultimi possono essere lipidi, proteine, carboidrati o combinazioni di tali macromolecole come glicoproteine e glicolipidi.
Alcuni farmaci antivirali agiscono proprio sugli anti-recettori dei virus, impedendone la penetrazione nella cellula.
In alcuni casi, tuttavia, il virus è dotato di un elevato tasso di mutazione e di variabilità delle molecole superficiali, vanificando di volta in volta i farmaci in utilizzo e imponendo una continua ricerca e messa a punto di agenti antivirali più efficaci.
Un esempio di tale problematica è la continua lotta nei confronti del virus HIV.
Anche i virus influenzali sono caratterizzati da una notevole variabilità, a causa della quale ogni anno vengono allestiti nuovi vaccini.
2) Penetrazione
La seconda fase è la penetrazione.
I virus animali entrano nella maggior parte dei casi mediante endocitosi o fusione con la membrana plasmatica.
In rari casi viene introdotto solo l'acido nucleico.
Generalmente l'envelope, se presente, viene perso a seguito della fusione con la membrana plasmatica, oppure eliminato in seguito all'intervento degli enzimi contenuti nei lisosomi con cui si fondono le vescicole endocitiche.
Il capside viene rimosso nel citoplasma o al contatto con la membrana nucleare.
Più complessa è la modalità di penetrazione dei batteriofagi. Essi aderiscono attraverso le spine caudali alla superficie batterica, le quali successivamente si contraggono e avvicinano la parte terminale della coda alla parete.
Un enzima crea un poro e a seguito della contrazione della coda l'acido nucleico viene iniettato nella cellula.
Nelle piante i virus vengono introdotti nelle cellule grazie alla proboscide degli insetti che si nutrono dei loro fluidi.
I virus animali senza enevelope sfruttano nella maggior parte dei casi il fenomeno dell'endocitosi per penetrare nella cellula ospite (1). In rari casi viene iniettato solo l'acido nucleico (2).
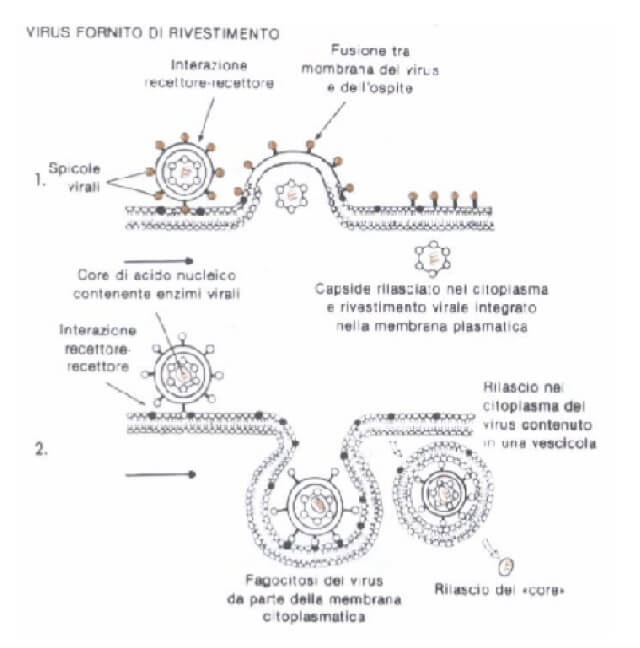
I virus animali con envelope entrano nella cellula ospite a seguito della fusione con la membrana plasmatica liberando nel citoplasma il nucleocapside (1) o mediante endocitosi (2).
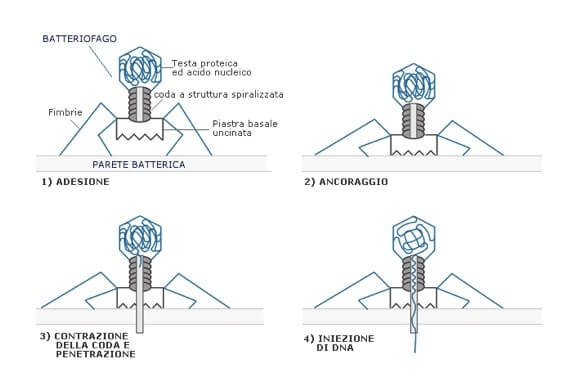
La penetrazione dei batteriofagi è tra le più complesse. Avvenuta l'adesione, le spine caudali si contraggono e avvicinano la coda alla parete. Un enzima perfora la parete, attraverso cui viene iniettato l'acido nucleico.
3) Replicazione: sintesi dell'acido nucleico e delle proteine
L'esposizione dell'acido nucleico virale agli enzimi cellulari avvia la fase replicativa in cui si producono numerose copie del genoma e vengono sintetizzate le proteine del capside.
Questa fase si differenzia tra i vari virus a seconda della tipologia dell'acido nucleico, della configurazione a doppio o a singolo filamento e della polarità dei genomi a RNA.
Le differenze riscontrabili consentono di classificare i virus in sette classi, secondo lo schema di Baltimore.
Caratteristiche sono le classi VI e VII per la presenza del fenomeno della trascrizione inversa, ossia la produzione di una molecola di DNA, detta DNA complementare (cDNA), da uno stampo a RNA.
Questo processo è realizzato da un enzima contenuto nel virione stesso, la trascrittasi inversa.
La classe VI è quella dei retrovirus, virus rivestiti con un genoma a RNA a polarità positiva, che provocano nell'uomo e negli animali alcune forme di tumore e comprendono il virus HIV.
Una volta avvenuta la penetrazione, il genoma a RNA viene retrotrascritto in DNA a doppio filamento che si integra nel genoma della cellula ospite.
Mediante regolare trascrizione vengono prodotti diversi RNA utilizzati sia come messaggeri per la sintesi proteica sia come genoma delle nuove particelle virali.
La classe VII comprende virus che hanno DNA a doppio filamento, duplicato attraverso un intermedio a RNA da una trascrittasi inversa.
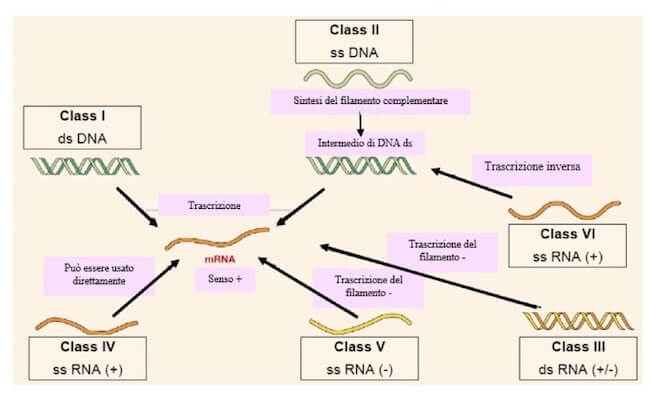
Le diverse modalità replicative dei virus a RNA e DNA. Le sigle ss e ds significano rispettivamente singolo e doppio filamento. I genomi a RNA + vengono usati direttamente come mRNA e per la duplicazione viene prima creato uno stampo -. I genomi a RNA – vengono duplicati da uno stampo + utilizzato anche come mRNA. La classe VI comprende i retrovirus, virus a RNA + da cui mediante trascrizione inversa viene creata una molecola di DNA ds (cDNA – DNA complementare) che si integra nel genoma della cellula ospite e trascritto in mRNA che sarà utilizzato sia per la sintesi proteica sia come genoma delle nuove particelle virali. Anche i virus della classe VII (non indicata) utilizzano la trascrittasi inversa.
4) Assemblaggio dei virioni
Nella quarta fase del ciclo replicativo virale vengono allestiti i capsidi ed eventualmente le componenti lipidiche degli envelope per creare nuovi virioni.
L'introduzione dell'acido nucleico al loro interno è detto impacchettamento.
5) Rilascio
Nel caso dei batteriofagi, i nuovi virioni assemblati nella cellula ospite vengono rilasciati in seguito a lisi e uccisione della cellula.
Anche i virus animali possono adottare questa strategia, ma nella maggior parte dei casi, soprattutto se rivestiti, fuoriescono per gemmazione.
Talvolta questo processo può essere lento e la cellula può rimanere viva per molto tempo dando luogo a un'infezione persistente.
Rilascio per gemmazione dei virioni nelle cellule animali. L'envelope origina dalla membrana plasmatica della cellula ospite, da cui si spiega la composizione chimica molto simile ad essa.
Il ciclo vitale dei batteriofagi può essere di tipo litico o lisogeno
Se un batteriofago, dopo la penetrazione nella cellula ospite, si indirizza subito vero la lisi della cellula si dice che realizza un ciclo litico.
Alcuni batteriofagi, invece, sono definiti temperati e realizzano un ciclo lisogeno, in cui il genoma virale, una volta introdotto nella cellula, si integra nel genoma batterico.
In tale stato il virus è detto profago e può rimanervi per molto tempo, duplicandosi insieme al genoma batterico durante la scissione binaria e perpetuandosi quindi in più cellule, senza che avvenga sintesi delle proteine virali.
Solo quando la sopravvivenza delle cellule è minacciata, ad esempio da esposizione a raggi ultravioletti, il genoma virale si separa da quello batterico e avvia il normale ciclo litico.
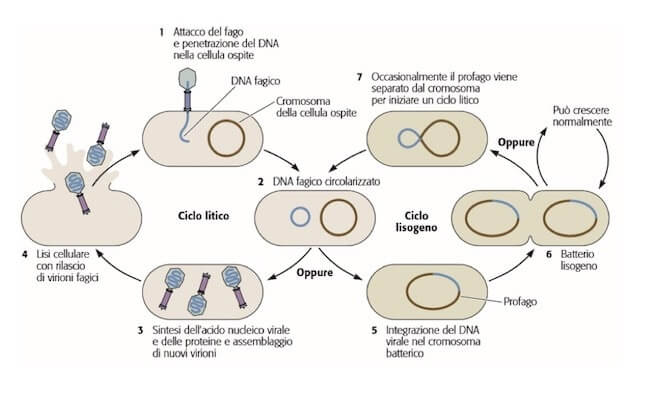
Il ciclo litico e il ciclo lisogeno dei batteriofagi.
Gli effetti dei virus sulle cellule animali
Gli esiti di un'infezione virale sulle cellule animali possono essere diversi.
Come detto precedentemente, alcuni danno infezioni litiche, causando la morte delle cellule ospiti.
Molti virus dotati di envelope fuoriescono per gemmazione.
Quando il rilascio dei nuovi virioni avviene lentamente e la cellula rimane vitale per lungo tempo si parla di infezione persistente.
Sono possibili anche delle infezioni latenti, se si verifica un ritardo tra il momento in cui avviene l'infezione e la lisi.
Un esempio è l'infezione da herpes simplex che causa l'erpete febbrile, i cui sintomi (la lisi delle cellule e l'escoriazione che ne deriva) compaiono sporadicamente quando il virus riemerge dalla latenza.
L'infezione latente provocata dai virus animali ricorda il ciclo lisogeno dei batteriofagi temperati, ma non prevede l'integrazione del genoma virale in quello della cellula ospite.
Infine, alcuni virus possono causare la trasformazione in cellula tumorale perché interferiscono con i meccanismi molecolari preposti al controllo del ciclo cellulare.
Tra i virus tumorigenici si ricordano alcuni retrovirus, responsabili di leucemie nell'uomo e negli animali, e tra i virus a DNA quelli dell'epatite B (HBV) e C (HCV), alcuni herpesvirus e papillomavirus.
Link correlati:
Qual è la differenza tra virus e batteri?
Che cosa sono i microrganismi patogeni?
Cosa sono le mutazioni geniche?
Che cosa sono le cellule natural killer?
Che cosa sono i provirus?
Virus difettivi: la transduzione
Che cosa sono i meccanorecettori?
Che cos'è la trasduzione?
Che cosa sono i trasposoni?
Che cosa sono i viroidi?
Che cosa sono gli infusori?
Studia con noi