Emoglobina
Struttura e funzioni dell'emoglobina
L'ossigeno, essendo poco solubile in acqua, non può essere trasportato a livello dei tessuti nel plasma sanguigno.
Per questo è necessario che l'ossigeno si leghi a qualche molecola nel torrente circolatorio.
I metalli di transizione, come il ferro e il rame, hanno una forte tendenza a legare l'ossigeno.
Il ferro metallico (stato di ossidazione 0) forma le specie reattive dell'ossigeno (ROS), che sono radicali liberi che possono danneggiare le macromolecole biologiche.
Invece, il ferro in forma ionica e precisamente nello stato di ossidazione +2, è meno reattivo del ferro metallico.
Esso è incorporato nel gruppo eme, un gruppo prostetico, cioè parte della molecola di una proteina coniugata non costituita da amminoacidi.
L'eme ha una struttura ad anello, la protoporfirina, legata a un atomo di ferro, nello stato di ossidazione +2.
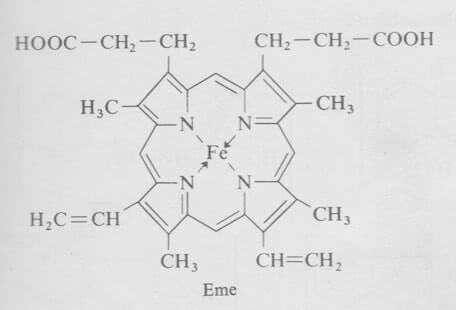
L'atomo di ferro può formare 6 legami di coordinazione: 4 con gli atomi di azoto facenti parte dell'anello porfirinico e altri due, perpendicolari al piano della proteina, uno dei quali lega un residuo di istidina (chiamata istidina prossimale) dell'emoglobina.
Gli atomi di azoto della protoporfirina, coordinati al ferro, fanno sì che esso non si ossidi allo stato +3.
Mentre il ferro nello stato +2 lega reversibilmente l'ossigeno, il ferro nello stato +3 non è in grado di legare il gas, formando la metaemoglobina, che mantiene la stessa struttura e può, invece, legare l'acqua.
Il sangue contiene i globuli rossi o eritrociti, i quali contengono l'emoglobina e quindi trasportano e rilasciano l'ossigeno.
Gli eritrociti normali nell'uomo hanno forma di disco biconcavo e il loro diametro va dai 6 ai 9 micron.
Durante il processo di maturazione a partire da cellule staminali, gli emocitoblasti, vengono prodotte le cellule figlie, che producono grandi quantità di emoglobina e perdono gli organelli citoplasmatici, come i mitocondri e il nucleo.
Non avendo il nucleo, i globuli rossi non possono replicarsi e sopravvivono per circa 120 giorni.
Nel sangue arterioso l'emoglobina è satura per circa il 96% di ossigeno, mentre in quello venoso l'emoglobina è saturata solamente per il 64%.
L'emoglobina è costituita da 4 subunità. Le interazioni tra queste determinano variazioni dell'affinità dell'emoglobina per l'ossigeno e questa modulazione di legame fa sì che la proteina possa rispondere in base alle richieste dei vari tessuti.
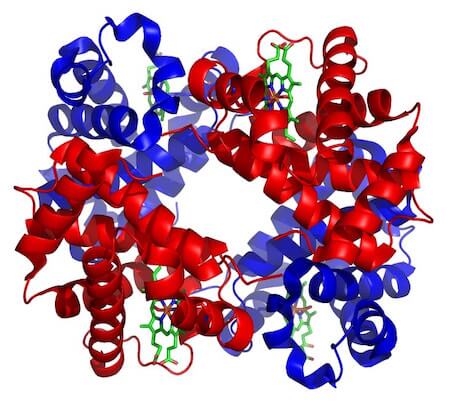
Struttura dell'emoglobina: le quattro subunità che la costituiscono sono rappresentate due in blu e due in rosso.
L'emoglobina ha una massa molare di 64500 g/mol, è una proteina globulare, quindi ha una forma pressoché sferica e un diametro di 5,5 nanometri. Ogni subunità è costituita da un gruppo eme.
L'emoglobina A è quella tipica degli adulti e contiene due tipi di globine, quindi le 4 subunità sono due catene α, costituite ognuna da 141 amminoacidi, e due catene β, costituite ognuna da 146 amminoacidi.
Le catene α e β hanno una struttura tridimensionale molto simile, pur avendo strutture primarie molto differenti.
La struttura tridimensionale di queste due subunità è simile a quella della mioglobina, che pure varia in quanto a struttura primaria.
Le 4 subunità dell'emoglobina formano interazioni molto forti tra loro.
L'interfaccia tra le subunità α1 e β1 e quella tra α2 e β2 sono costituite ognuna da circa 30 residui amminoacidici ed è abbastanza forte.
Le interfacce tra α1 e β2 e tra α2e β1, invece, comprendono circa 20 residui.
Nelle interfacce prevalgono le interazioni idrofobiche, ma sono presenti anche alcuni legami a idrogeno e coppie ioniche (o ponti salini).
Meccanismo dell'emoglobina
Dall'analisi ai raggi X, è stato possibile vedere che l'emoglobina è presente in due conformazioni: lo Stato R (rilassato) e lo Stato T (teso).
L'ossigeno ha una maggiore affinità per lo Stato R, che viene stabilizzato dall'interazione con il gas, mentre, in assenza di ossigeno, lo stato T è quello più stabile ed è la conformazione della cosiddetta deossiemoglobina.
L'emoglobina deve legare in maniera efficiente l'ossigeno nei polmoni, in cui la pressione parziale di ossigeno è di circa 13,3 kPa (kilopascal), e rilasciare l'ossigeno nei tessuti, dove la pressione è di 4 kPa.
L'emoglobina, passando dallo stato a bassa affinità per l'ossigeno a quello R, è la proteina ideale per portare l'ossigeno ai tessuti.
La prima molecola di ossigeno si lega ad una subunità dell'emoglobina, quindi la prima molecola si lega debolmente, perché la subunità è nello stato T.
In questo modo essa subisce una modificazione conformazionale che influenza anche le subunità adiacenti, facilitando quindi l'interazione delle altre subunità con l'ossigeno.
Ogni subunità lega una molecola di ossigeno, quindi una molecola di emoglobina lega 4 molecole di ossigeno e il legame della prima facilita il legame delle altre.
Questo tipo di legame è chiamato legame cooperativo.
Quindi, l'emoglobina è una proteina allosterica. Questa parola deriva dal greco allo (altro) e stereo (forma).
In una proteina allosterica, il legame del ligando al sito modifica le proprietà di un altro sito sulla stessa molecola proteica.
Quando il normale ligando di una proteina allosterica è anche un modulatore, come in questo caso, si parla di interazione omotropica.
Il fatto che l'emoglobina sia una proteina allosterica fa sì che essa possa rispondere in maniera più sensibile alle variazioni nella concentrazione di ossigeno.
Il legame cooperativo dell'ossigeno all'emoglobina venne scoperto nel 1910 da Hill.
Regolazione dell'emoglobina
L'emoglobina può trasportare anche protoni (H+) e anidride carbonica (CO2), due prodotti finali della respirazione cellulare dai tessuti verso i polmoni e i reni, dove queste sostanze sono escrete.
L'anidride carbonica prodotta dalla respirazione mitocondriale in seguito all'ossidazione delle molecole organiche, subisce reazione di idratazione, formando in maniera reversibile protoni e ioni bicarbonato.
Questa reazione è catalizzata dall'enzima anidrasi carbonica, presente in grandi quantità nei globuli rossi.
L'anidride carbonica è poco solubile in acqua e quindi deve essere convertita in bicarbonato (HCO3−).
La formazione di ioni H+ determina anche una riduzione del pH nei tessuti e si è visto che il legame dell'ossigeno è sensibile sia al pH che alla concentrazione di anidride carbonica.
Il legame di queste due molecole è inversamente proporzionale a quello dell'ossigeno all'emoglobina.
Nei tessuti periferici, in cui il pH è basso (elevata quantità di protoni), la concentrazione di anidride carbonica è alta, l'affinità dell'emoglobina per l'ossigeno è ridotta e il gas è rilasciato nei tessuti.
Nei polmoni, invece, l'anidride carbonica viene espulsa e, quindi, si trova presente in scarsa quantità.
Ciò fa sì che l'affinità dell'ossigeno sia elevata e il gas possa legarsi all'emoglobina per essere trasportato ai tessuti.
L'azione esercitata da protoni e anidride carbonica sull'affinità dell'ossigeno per l'emoglobina viene chiamata effetto Bohr.
La funzione dell'emoglobina è regolata anche dal 2,3-bisfosfoglicerato (BPG), il quale è presente in alte concentrazioni nei globuli rossi.
L'affinità dell'emoglobina per l'ossigeno è minore quando la concentrazione di BPG aumenta.
Esso si lega in un sito diverso da quello per l'ossigeno e modifica l'affinità di legame dell'ossigeno all'emoglobina in base alla pressione di ossigeno (pO2) nei polmoni.
L'ossigeno trasferito ai tessuti è circa il 40% della quantità di gas che può essere trasportata nel sangue, a livello del mare.
Invece, ad alta quota, la pressione di ossigeno è molto più bassa e il trasferimento di ossigeno ai tessuti si riduce.
Dopo poche ore, la concentrazione di BPG nel sangue aumenta e causa una riduzione dell'affinità dell'emoglobina per l'ossigeno e il trasferimento di ossigeno ai tessuti torna ad essere a livello fisiologico.
Le persone che soffrono di ipossia (ridotta ossigenazione dei tessuti periferici per problemi polmonari o cardiovascolari) presentano maggiori quantità di BPG.
I feti hanno un'emoglobina differente.
Al posto delle subunità β, sono presenti subunità γ.
L'emoglobina fetale, che ha un'affinità molto bassa per il BPG, presenta un'elevata affinità per l'ossigeno.
Questo è dovuto al fatto che il feto ottiene l'ossigeno dal sangue della madre, quindi è necessario che la sua emoglobina abbia una maggiore affinità rispetto a quella materna.
Carbossiemoglobina
Il complesso stabile emoglobina-monossido di carbonio prende il nome di carbossiemoglobina.
Anemia falciforme
Sono note almeno 500 varianti genetiche dell'emoglobina umana, di cui alcune molto rare.
Molte differiscono per un solo amminoacido, quindi subiscono mutazioni puntiformi.
Spesso queste non causano gravi conseguenze, ma, in alcuni casi, possono determinare serie patologie.
L'anemia falciforme colpisce soggetti che ereditano l'allele dell'emoglobina a cellule falciformi da entrambi i genitori.
Il numero di eritrociti si riduce e la loro forma è alterata: essi hanno forma allungata (a forma di falce).
L'emoglobina normale, chiamata emoglobina A, è molto più solubile di questo tipo di emoglobina, l'emoglobina S.
Le fibre insolubili dell'emoglobina S sono la causa della deformazione dei globuli rossi.
L'emoglobina S contiene un residuo di valina anziché un residuo di glutammato nelle due catene β.
La valina non ha la carica elettrica negativa che possiede il glutammato in catena laterale, quindi forma un'interazione idrofobica, che porta a un'anomalia strutturale della proteina.
Coloro che ricevono l'allele da uno solo dei genitori (eterozigoti) presentano una forma della malattia meno grave, che prende il nome di tratto dell'anemia falciforme, in cui solo l'1% degli eritrociti diventa falciforme.
Chi soffre di anemia falciforme presenta i seguenti sintomi: debolezza, respiro corto, problemi cardiaci, tachicardia e anche stati confusionali.
Essi presentano solo la metà di emoglobina normale perché le cellule sono fragili e si rompono facilmente.
Si verifica quindi una condizione di anemia (che letteralmente vuol dire mancanza di sangue).
Le cellule falciformi possono intasare i capillari dei tessuti a causa della loro forma, portando a dolore e malfunzionamento dell'organo colpito.
È questo a determinare la morte prematura dei soggetti con anemia falciforme.
La frequenza dell'allele per l'anemia falciforme è particolarmente elevata in alcune zone dell'Africa e del Mediterraneo, poiché l'allele conferisce una certa resistenza verso forme di malaria letali.
La malaria è causata dal parassita protozoo Plasmodium, che, inoculato dalla puntura della zanzara Anopheles, penetra nei globuli rossi più piccoli, spessi e deformati.
All'interno di questi, il parassita non è in grado di riprodursi, per questo, nelle zone malariche, coloro che sono sopravvissuti alla malattia sono prevalentemente le persone portatrici di anemia.
Quindi la selezione naturale ha portato alla diffusione di una popolazione anemica.
Analisi del sangue dell'emoglobina
I valori normali di emoglobina sono: 14-18 g/dl (per gli uomini); 12-16 g/dl (per le donne).
Valori di emoglobina al di sotto di queste soglie sono correlati prevalentemente ad anemia, carenza di ferro o gravidanza. Meno frequentemente, bassi livelli di emoglobina sono indice di patologie tumorali, come la leucemia, o di avvelenamento da piombo.
Per contrastare la carenza di emoglobina, nei casi più gravi, vengono effettuate trasfusioni di sangue.
Valori di emoglobina superiori a quelli medi, invece, possono essere rilevati in caso di shock, enfisema, ustione, disidratazione o a seguito di ripetute trasfusioni di sangue.
Le aptoglobine costituiscono un gruppo di proteine del siero del sangue caratterizzate dalla capacità di legare l'emoglobina.
Link correlati:
Vuoi sapere perché il sangue è rosso?
Che cos'è l'ossiemoglobina?
Cosa sono le mutazioni geniche?
Studia con noi