Ciclo di Cori
Che cos'è il il ciclo di Cori?
Il metabolismo energetico delle nostre cellule è basato sull'utilizzo dei carboidrati, il glucosio in particolare, come fonte per la produzione di ATP (la moneta energetica della vita) al termine del processo ossidativo completo del glucosio, appunto, ad anidride carbonica ed acqua, con una serie di reazioni chimiche che nel loro insieme costituiscono la respirazione cellulare.
Il metabolismo energetico, ovvero tutte le vie cataboliche che hanno lo scopo di produrre energia, convergono nel ciclo degli acidi tricarbossilici (ciclo di Krebs), quindi nella catena di trasporto degli elettroni e nella fosforilazione ossidativa ove, l'ATPsintetasi, fosforila l'ADP ad ATP utilizzando un fosfato inorganico e l'energia derivante dal catabolismo ossidativo.
La fonte principale, come detto, di energia per le nostre cellule è rappresentata dai carboidrati ma esistono anche vie metaboliche che degradano rispettivamente le proteine e i trigliceridi per la produzione di ATP (vedi schema nella figura seguente).
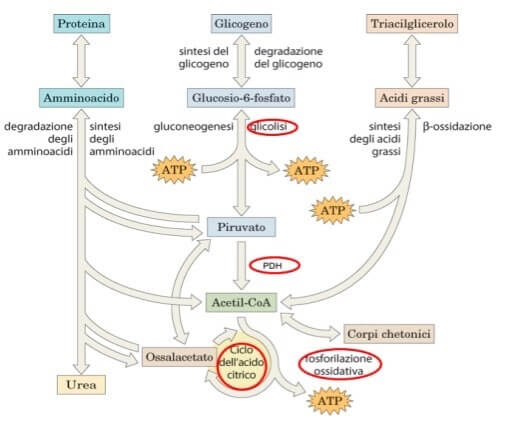
Metabolismo energetico delle cellule umane: le principali vie. In rosso sono evidenziate le reazioni della respirazione cellulare.
Ogni tessuto del nostro corpo umano presenta funzioni diverse e specifiche che si riflettono sia nella sua anatomia ed istologia, sia nella sua attività metabolica.
Ad esempio, le cellule nervose, i neuroni, utilizzano prevalentemente glucosio.
Questo ci conduce alla centralità di un organo che può essere definito la “centrale chimica” del corpo, ovvero il fegato (ti potrebbe interessare: dove si trova il fegato).
È infatti compito del fegato liberare glucosio nel circolo sanguigno affinché organi e tessuti, come il sistema nervoso centrale (SNC) appunto, possano rifornirsene.
Il fegato assolve a questa funzione sia liberando glucosio dalle riserve epatiche di glicogeno, sia attraverso la gluconeogenesi (l'anabolismo di glucosio a partire da metabolici gluconeogenici ma non di derivazione glucidica).
Per indicare la centralità metabolica degli epatociti spesso si utilizza anche il termine di “organi extraepatici” per indicare tutti gli altri organi periferici del nostro corpo che dipendono in qualche misura dal metabolismo epatico per i precursori delle vie metaboliche che li caratterizzano.
Dopo essere stati assorbiti a livello intestinale, infatti, carboidrati (monosaccaridi), amminoacidi e triacilgliceroli (trigliceridi) passano nel sangue tramite la vena porta e raggiungono gli epatociti.
Essi trasformano le sostanze nutrienti in composti che possono generare energia e in precursori necessari agli altri tessuti.
I precursori metabolici vengono inviati alla loro destinazione finale attraverso il flusso sanguigno.
I tipi e le quantità di sostanze nutrienti e precursori prodotti dagli epatociti variano in base a diversi fattori tra cui la composizione della dieta e l'intervallo di tempo tra un pasto e l'altro.
La richiesta di energia e di precursori dei tessuti extraepatici varia allo stesso modo da organo a organo sulla base dell'attività dell'organismo.
La sintesi dei precursori e dei nutrienti a livello epatico è regolata da ormoni e dalla presenza di enzimi allosterici.
Un tessuto che necessita di molta energia, come il tessuto muscolare scheletrico riflette questa necessità nel suo metabolismo, infatti le fibrocellule muscolari striate hanno un metabolismo del glucosio ad uso esclusivo e, diversamente da tutte le altre cellule periferiche, scorte proprie di glicogeno.
In condizioni di riposo e dopo i pasti, il muscolo sintetizza e accumula glicogeno, che arriva a costituire l'1-2% della sua massa.
Il glicogeno è trasformato in glucosio-6-fosfato per entrare nella glicolisi e dunque nel metabolismo ossidativo per la sintesi di ATP, ma non può essere esportato perché manca la glucoso-6-fosfatasi.
Il glucosio ematico è non fosforilato, mentre quando entra nelle cellule viene fosforilato in posizione 6 in modo da non poter lasciare la cellula stessa, non inficiare il gradiente tra interno ed esterno della cellula e per essere indirizzato alle varie vie metaboliche.
Solo il fegato può idrolizzare il fosfato del glucosi ad opera della glucosio-6-fosfatasi ed immettere quindi il monosaccaride nel circolo ematico, tutte le altre cellule, incluse le fibrocellule muscolari striate non possono liberare glucosio nel sangue.
Nel muscolo inoltre è attiva la glicogeno sintesi tramite la glicogeno sintetasi per la produzione di glicogeno muscolare di scorta, mentre non è attiva la gluconeogenesi perché non sono espressi gli enzimi necessari, presenti invece a livello epatico.
Nel muscolo il metabolismo dei carboidrati è perciò a uso esclusivo di questo tessuto.
Reazioni e significato del Ciclo di Cori
Nelle fibrocellule muscolari striate in condizione di aerobiosi (concentrazione fisiologica di ossigeno ematico e tissutale) il glocosio-6-fosfato è indirizzato alla glicolisi e si giunge alla produzione del piruvato.
In queste condizioni il destino del piruvato è ossidativo, ovvero tramite la piruvato deidrogenasi (PDH) viene convertito, nella matrice mitocondriale in acetil-CoA per essere poi degradato nel ciclo di Krebs e portare a termine la respirazione cellulare con al catena di trasporto degli elettroni e la fosforilazione ossidativa.
In condizioni di anaerobiosi (maggior sforzo fisico e minore apporto di ossigeno) il destino del piruvato è differente ed esso viene riconvertito in un metabolita ad uno stadio ossidativo minore, l'acido lattico, nella fermentazione lattica.
Essa ha lo scopo di riconvertire NAD ridotto in NAD+ ossidato al fine di rifornire la glicolisi che diversamente verrebbe rallentata.
Se non è, infatti, disponibile l'ossigeno, nelle nostre cellule, non può avvenire la fosforilazione ossidativa e non viene prodotto ATP, i muscoli dunque possono utilizzare solo le 2 molecole di ATP prodotte per fosforilazione a livello di substrato della glicolisi.
Se non avviene la fosforilazione ossidativa i NAD ridotti non sono ossidati e dunque non sono disponibili per la gliceraldeide fosfato deidrogenasi in glicolisi, portando al rallentamento della via metabolica.
Con la fermentazione lattica si consente, da un lato di non rallentare la glicolisi permettendo la continua sintesi di 2 ATP nel citosol e dall'altro di convertire il piruvato (un acido) in un metabolita come il lattato anch'esso acido (acido lattico) e che, dunque, darebbe problemi al pH cellulare ma che può essere rilasciato nel circolo ematico.
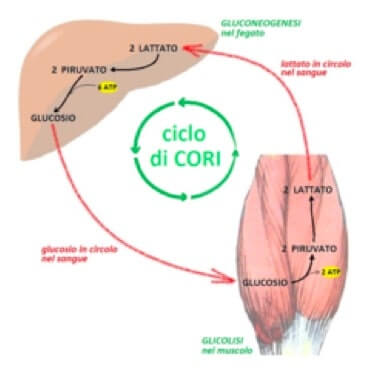
Il ciclo di Cori: attraverso il circolo sanguigno il lattato prodotto nel muscolo raggiunge il fegato, ove è riconvertito il glucosio. Il glucosio rilasciato dagli epatociti nel sangue, raggiungerà il muscolo per la produzione di energia.
Un'altra via tramite la quale il piruvato può essere allontanato dal nucleo è tramite transaminazione a produrre alanina.
Questa, tramite il circolo sanguigno, raggiunge il fegato che la riconverte in piruvato, elimina l'NH2 attraverso il ciclo dell'urea e converte il piruvato in glucosio con la gluconeogenesi. In questo modo il muscolo smaltisce l'azoto in eccesso.
Il passaggio di acido lattico e di alanina, tramite la circolazione ematica, tra fibrocellule muscolari striate e epatociti, prende il nome di ciclo di Cori e ciclo glucosio-alanina e sono un ottimo esempio di integrazione metabolica tra tessuti ed organi diversi, esemplificativa della complessità ma anche della perfezione della fisiologia del nostro corpo.
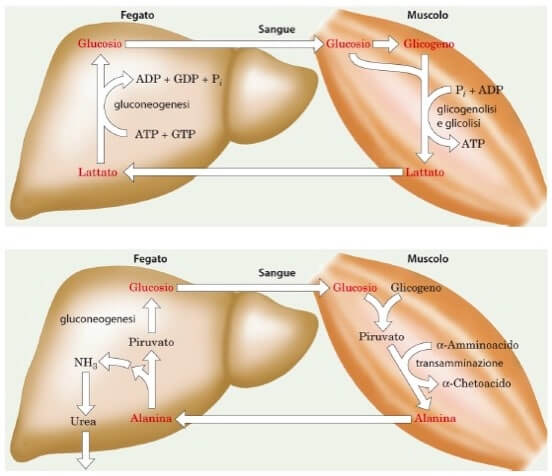
Il Ciclo di Cori (in alto) e il Ciclo glucosio-alanina (in basso): due esempi di metabolismo integrato.
Gli altri tessuti utilizzano altri trasportatori per l'azoto in eccesso che deve comunque raggiungere gli epatociti per accedere al ciclo dell'urea ed essere catabolizzato correttamente ed eliminato tramite la filtrazione renale.
Si tratta dell'acido aspartico e dell'acido glutammico che, grazie alla transaminazione si formano rispettivamente dall'ossalacetato e dall'α-chetoglutarato.
L'accettore finale di gruppi amminici, nel fegato, è l'α-chetoglutarato che si trasforma in glutammato.
Link correlati:
Quali sono le otto reazioni del ciclo dell'acido citrico?
Che cosa sono i cicli biogeochimici?
Che cosa è il ciclo cellulare?
Studia con noi