Bottone sinaptico
Che cos'è il bottone sinaptico?
Il termine sinapsi deriva dal greco e significa "collegamento/congiungere", in neurofisiologia si intende il contatto tra due cellule eccitabili, in particolare definisce il punto di contatto funzionale tra le due cellule (due cellule nervose, oppure una cellula nervosa ed una fibrocellula muscolare) che permette il passaggio dell'eccitazione da una all'altra in una sola direzione. Si classificano due tipologie di sinapsi: la sinapsi elettrica e la sinapsi chimica.
La sinapsi elettrica è un punto di contatto tra le due membrane cellulari delle due cellule coinvolte, grazie al quale il potenziale, l'impulso elettrico di una cellula viene propagato alla cellula successiva. La membrana e la cellula che trasmette l’impulso è definita pre-sinaptica, quella ricevente post-sinaptica.
Gli eventi elettrici che generano e trasmettono l'impulso tra le due cellule, sono veicolati da speciali proteine con la funzione di canali ionici e recettori canale. L'impulso elettrico viene generato grazie a cambiamenti conformazionali in tali recettori e/o canali e dal conseguente trasferimento di ioni carichi (Na+, K+, Cl- e Ca2+ principalmente) dal un lato all'altro della membrana plasmatica ovvero dall’interno all’esterno delle cellule e viceversa.
Che cos'è e quali funzioni svolge il bottone sinaptico?
Nelle sinapsi chimiche si definisce bottone sinaptico il complesso che include, la membrana della cellula pre-sinaptica, la membrana della cellula post-sinaptica lo spazio tra le due.
In questo caso l'impulso elettrico, il potenziale d'azione, generato dalla cellula pre-sinaptica, giunge al bottone sinaptico e provoca la fuoriuscita del cosiddetto neurotrasmettitore ovvero una molecola chimica.
Quest'ultimo verrà liberato nello spazio sinaptico ed andrà a legarsi a speciali recettori presenti nella membrana post-sinaptica dove provocherà dei cambiamenti a livello di ioni tali per cui si genererà un cambiamento nel potenziale di membrana (con generazione di un potenziale d'azione) e dunque l'impulso sarà trasmesso.
Nelle sinapsi chimiche quindi l’informazione nasce come elettrica, viene tramutata in segnale chimico e poi nuovamente in elettrico, mentre nelle sinapsi elettriche il segnale è sempre elettrico.
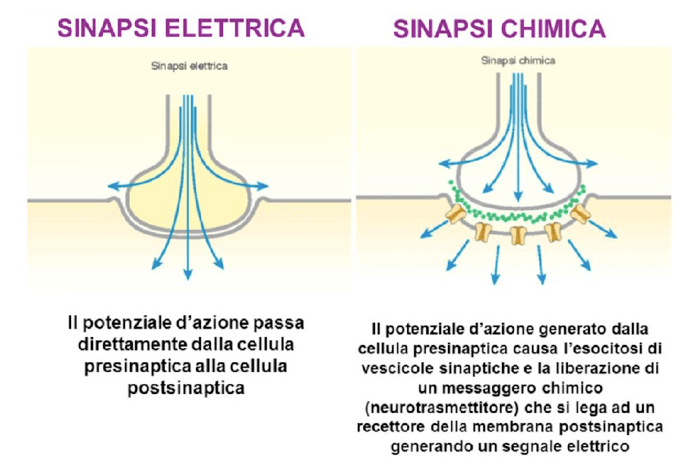
I due tipi di sinapsi: elettrica e chimica.
Abbiamo già ricordato che la trasmissione dell'impulso è unidirezionale, dalla cellula che lo genera a quella che lo riceve.
Morfologia del neurone
Ricordiamo ora la morfologia della tipica cellula nervosa, ovvero il neurone. Il neurone è costituito da un corpo cellulare, detto soma nel quale si trovano il nucleo e gli organelli, di un lungo prolungamento detto assone che termina con le sinapsi e rappresenta quindi il terminale pre-sinaptico del collegamento che quel neurone instaura con la cellula (sia essa un altro neurone oppure una fibrocellula muscolare) ricevente.
Il potenziale d'azione si genera nel cosiddetto cono, ovvero il punto del soma dove nasce l'assone e si propaga per tutta la sua lunghezza fino alla sinapsi.
Il potenziale d'azione viaggia grazie ad una modalità definita saltatoria, ovvero non percorre tutta la lunghezza della membrana dell'assone ma salta da un punto ad un altro. I punti in cui il potenziale d'azione può saltare sono le porzioni della membrana plasmatica non rivestiti da mielina, definiti nodi di Ranvier. Infatti, l'assone, per tutta la sua lunghezza è rivestito dalla mielina, cioè dalla membrana plasmatica di speciali cellule accessorie, gli oligodendrociti a livello centrale e le cellule di Schwann a livello periferico, che lo avvolgono più volte formando una sorta di manicotto.
Nel neurone sono inoltre presenti prolungamenti più brevi dell'assone e di solito presenti un numero molto maggiore, definiti dendriti. Tramite i dendriti le cellule nervose ricevono le informazioni dagli altri neuroni a formare una rete di trasmissione di impulsi. Ogni neurone, infatti, può ricevere più afferenze da più neuroni che, coi loro rispettivi assoni, lo contattano a livello del soma oppure dei dendriti e genera a sua volta un potenziale d'azione che tramite il suo lungo assone viene "scaricato" alla cellula efferente.
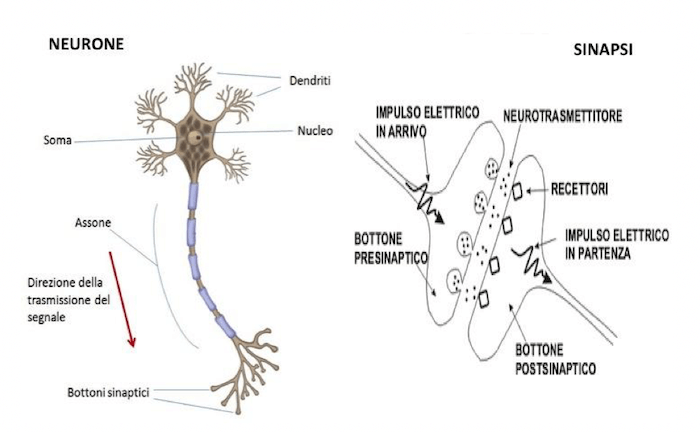
Schema raffigurante un tipico neurone e una sinapsi chimica. Il segnale si trasmette in maniera unidirezionale, il neurone riceve impulsi tramite il soma e i dendriti e li trasmette tramite l'assone.
La trasmissione di tipo saltatorio del potenziale d'azione negli assoni, permette di raggiungere velocità maggiori e, come abbiamo già detto, è permessa dalla mielina.
Le cellule responsabili della mielina sono cellule appartenenti alla cosiddetta macroglia, si tratta di cellule compagne dei neuroni, che non generano e trasmettono direttamente gli impulsi elettrici ma costituiscono con i neuroni delle unità funzionali.
Appartengono alla macroglia:
- gli astrociti responsabili del sostegno metabolico dei neuroni e della formazione della barriera ematoencefalica,
- gli oligodendrociti responsabili dalla mielina a livello centrale
- le cellule di Schwann responsabili della mielina a livello periferico.
A livello centrale un singolo oligodendrocita riveste diversi assoni di diversi neuroni, mentre, a livello periferico, un'unica cellula di Schwann riveste un solo assone.
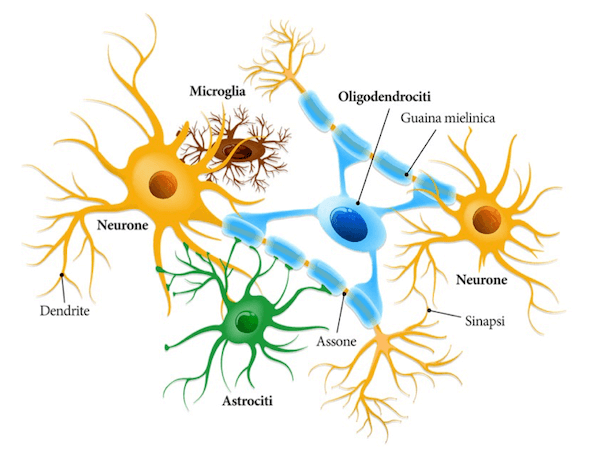
Lo schema mostra la formazione della mielina da parte di un oligodendrocita che riveste gli assoni di più neuroni ed una rappresentazione di astrociti e microglia.
Oltre alla macroglia, detta anche semplicemente glia, nel sistema nervoso si trova la cosiddetta microglia rappresentata da cellule con una diversa origine embrionale; se la macroglia, infatti, ha la stessa origine dei neuroni ovvero dal foglietto ectodermico, dal neuroectoderma in particolare, la microglia è di origine immunitaria e dunque origina embriologicamente dal mesoderma.
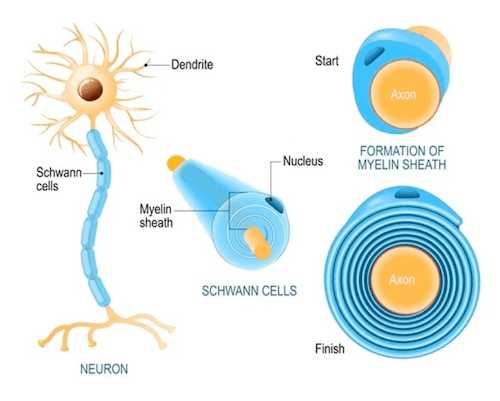
Formazione della mielina da parte di una cellula di Schwann.
Il potenziale d'azione
Il potenziale d'azione è l'impulso elettrico di cui abbiamo parlato in precedenza, in grado di propagarsi tra neuroni oppure tra un neurone ed una cellula muscolare. Si tratta di un cambiamento del potenziale di membrana con delle ben precise caratteristiche identificate grazie agli studi di elettrofisiologia su cellule eccitabili.
Prima di definire meglio il potenziale d'azione, è necessario capire che cosa si intende per potenziale di membrana, ovvero la differenza di potenziale elettrico, misurabile grazie a microelettrodi inseribili nelle cellule, che si insatura dai due lati delle membrane plasmatiche: il lato intracitoplasmatico ed il lato extracitoplasmatico.
Questo accade poiché all'interno della cellula sono presenti ioni ed altre molecole cariche in concentrazione diversa rispetto all'ambiente extracellulare. Ogni cellula del nostro corpo deve raggiungere l'equilibrio osmotico cioè la situazione nella quale essa presenta una distribuzione differenziale di ioni ai capi della membrana obbedendo ai principi elettrochimici.
Questi principi stabiliscono che tutte le molecole tendono a distribuirsi uniformemente tra due compartimenti separati da un setto che sia attraversabile dalle molecole, come accade ad una goccia di inchiostro che, se posta all'interno di un bicchiere d'acqua tenderà a distribuirsi uniformemente in tutto il volume (questo in chimica è definito come movimento secondo gradiente).
Le molecole si muovono continuamente con movimenti casuali ma se esiste una differenza di concentrazione tra due punti, dunque se esiste quello che si definisce un gradiente (come nel bicchiere e dentro la goccia dove l'inchiostro è più concentrato) si instaurerà un movimento netto di molecole da dove esse sono maggiormente concentrate a dove lo sono meno, fino al raggiungimento dell'equilibrio, momento nel quale non si noterà più un movimento netto ma continueranno i movimenti casuali.
Il movimento secondo gradiente in assenza di ostacoli, come nel bicchiere con la goccia di inchiostro del nostro esempio, si definisce diffusione semplice.

Esemplificazione del movimento secondo gradiente definito diffusione semplice.
Le nostre cellule, però, sono racchiuse dalla membrana cellulare, la membrana plasmatica costituita da un doppio strato di fosfolipidi che è definita una membrana semipermeabile poiché lascia passare, per diffusione semplice, alcune molecole mentre è impermeabile ad altre, in particolare attraversano per diffusione semplice le membrane plasmatiche le piccole molecole di gas come l'ossigeno e l'anidride carbonica, le molecole idrofobiche in grado di "sciogliersi" nel doppio strato fosfolipidico, mentre non sono in grado di attraversare le membrane plasmatiche le molecole cariche come gli ioni e le molecole polari come il glucosio. L'acqua, pur essendo una molecola polare, riesce ad attraversare il doppio strato fosfolipidico in virtù delle sue dimensioni.
Gli ioni e le molecole polari attraversano le membrane plasmatiche solo grazie a speciali molecole trasportatrici come i canali oppure le pompe come ad esempio la pompa sodio potassio, tale trasporto potrà avvenire in maniera passiva, secondo gradiente oppure contro gradiente con dispendio di energia, quindi la cellula, il trasportatore nello specifico, consumerà energia, di solito molecole di ATP (adenosina trifosfato) per compiere il lavoro di trasporto.
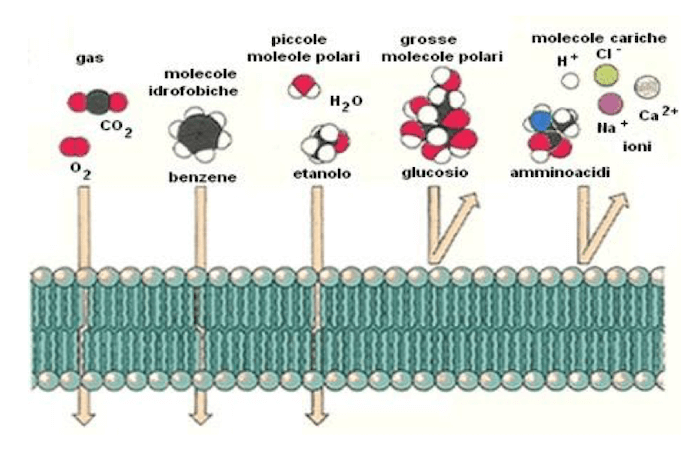
Lo schema mostra quali tipologie di molecole sono in grado di attraversare, per diffusione semplice, la membrana plasmatica delle nostre cellule.
Oltre al gradiente chimico gli ioni rispondono ad un principio elettrico per cui gli anioni sono attratti da cariche positive e i cationi da quelle negative dunque uno ione tenderà a disporsi ai due lati della membrana plasmatica cellulare in virtù sia della sua concentrazione che della distribuzione di cariche ai lati della membrana stessa, ovvero tra l'interno e l'esterno della cellula.
All'interno di tutte le cellule sono presenti molecole proteiche che sono responsabili di una distribuzione diversa di cariche ai due lati della membrana plasmatica, in particolare l'interno delle cellule è maggiormente negativo rispetto all'esterno. Di conseguenza esiste una differenza di potenziale elettrico ai due lati della membrana plasmatica.
Esistono nel nostro corpo cellule definite eccitabili, ovvero in grado di modulare il potenziale di membrana in risposta a stimoli provenienti dall'ambiente e dunque di generare risposte elettriche che possono fungere da segnale. Le cellule eccitabili per eccellenza del nostro corpo, sono senza dubbio i neuroni. Queste cellule ricevono segnali elettrici, analizzano l'informazione e generano una risposta appropriata.
Il segnale fondamentale, atto a trasmettere l'informazione, è il cosiddetto potenziale d'azione, un segnale elettrico che può essere trasmesso lungo le membrane degli assoni dei neuroni, sia quelli sensitivi che motori.
Nelle cellule in grado di eccitarsi, la permeabilità ionica della membrana plasmatica può variare, non è fissa come per le altre cellule ed è proprio questo che genera il segnale elettrico. In particolare è l'aumento transitorio della permeabilità al sodio alla base della creazione del potenziale d'azione, ovvero del segnale elettrico di base del nostro sistema nervoso.
Il potenziale di membrana, a riposo, di un neurone è stato misurato con particolari microelettrodi ed ha un valore di –70 mV. Il potenziale d'azione è generato dall'apertura di canali per il sodio che dunque rendono la membrana permeabile a questo ione. L'ingresso di ioni sodio, positivi, provoca quella che prende il nome di depolarizzazione di membrana, ovvero i valori del potenziale di membrana diventano meno negativi, per tornare poi, successivamente, a valori normali.
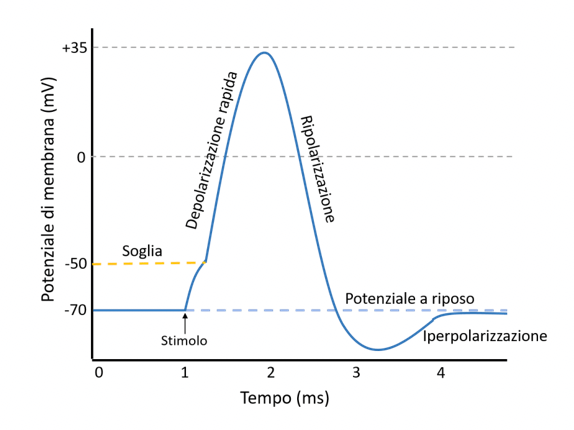
Grafico del potenziale d'azione: esso si genera solo superata una certa soglia di valore di potenziale di membrana ed è caratterizzato da una prima depolarizzazione cui segue una bipolarizzazione, una breve fase di iperpolarizzazione e dunque il ritorno al valore del potenziale di membrana a riposo.
Il potenziale d'azione ha caratteristiche proprie:
- È evocato dal fatto che lo stimolo esterno provoca la depolarizzazione della membrana ma, per evocare il potenziale d'azione, la depolarizzazione deve superare un valore di soglia, dell'ordine di 10-20 mV. Quindi, dato il potenziale a riposo di –70 mV, per generare il potenziale d'azione, il potenziale di membrana deve giungere al valore di –60-50 mV.
- L'ampiezza del potenziale d'azione è indipendente dalla grandezza dello stimolo, ovvero, se lo stimolo è abbastanza grande da superare la soglia, il potenziale viene evocato con ampiezza e forma costante (come da grafico riportato in figura).
- La propagazione del potenziale d'azione lungo il neurone avviene senza attenuazioni, la velocità di conduzione del potenziale d'azione nelle fibre nervose è di circa 10-20 m/s (ma sono state osservate velocità fino a 100 m/s).
- Al picco del potenziale d'azione si verifica un'inversione di polarità del potenziale di membrana, ovvero l'interno diviene più positivo dell'esterno, questa è la fase del potenziale postumo ripolarizzante.
- Al termine del potenziale d'azione, l'assone si trova in uno stato definito di refrattarietà assoluta, il che significa che non è assolutamente in grado di generare un altro potenziale d'azione.
Quest'ultima caratteristica pone un limite alla frequenza possibile per il potenziale d'azione, infatti, poiché il periodo di refrattarietà è generalmente di circa 1 ms, ne consegue che non è possibile, per un neurone, generare più di 1000 potenziali d'azione al secondo.
La propagazione del potenziale d'azione
I ricercatori hanno scoperto, grazie a diversi esperimenti di elettrostimolazione, come l'apertura e la chiusura di speciali canali per il sodio e per il potassio, sia in grado di generare il potenziale d'azione; allo stesso modo è possibile spiegarne la propagazione lungo la membrana dell'assone del neurone.
Una volta generato il potenziale d'azione in un punto, la forte depolarizzazione indotta nelle regioni circostanti produce l'apertura dei canali del sodio e, quindi, una depolarizzazione rigenerativa che si trasmette via via nelle regioni adiacenti. Così si giustifica il fatto che il potenziale d'azione si trasmetta invariato in forma ed ampiezza lungo la fibra nervosa.
Esso può essere interpretato come un'onda depolarizzante che si autopropaga lungo la fibra nervosa perché ogni segmento successivo di membrana risponde alla depolarizzazione del segmento precedente.
I fisiologi hanno scoperto che la propagazione lungo gli assoni è sperimentalmente più rapida rispetto a quanto si possa teorizzare grazie al fenomeno appena discusso e questo perché in verità in molti neuroni quello che si verifica è un andamento saltatorio del potenziale d'azione. Ovvero, il segnale elettrico non si propaga fisicamente per tutta la lunghezza della membrana assonica ma salta da un Nodo di Ranvier all'altro, dunque si muove ad una velocità ancora maggiore. Ricordiamo che i cosiddetti Nodi di Ranvier sono le regioni assoniche non circondate da mielina, ovvero che restano scoperte.
Nel nostro SNC (sistema nervoso centrale) sono presenti sia fibre nervose mielinizzate che amielinizzate, in queste ultime la propagazione del potenziale d'azione è più lenta rispetto alle prime.
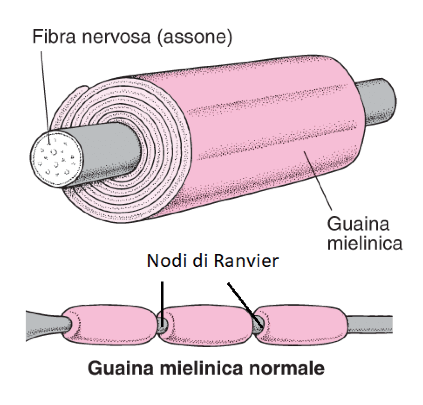
La figura mostra una porzione di un assone di una fibra nervosa mielinizzata e i Nodi di Ranvier, ovvero le regioni scoperte dell'assone.
Link correlati:
Che cosa sono i lisosomi?
Quanti neuroni abbiamo nel cervello?
Studia con noi