Eucromatina ed eterocromatina
Che cosa sono l'eucromatina e l'eterocromatina?
I termini eucromatina ed eterocromatina si riferiscono a due diversi aspetti che la cromatina assume nel nucleo delle cellule eucariotiche: la porzione di cromatina trascrizionalmente attiva e meno condensata si chiama eucromatina, mentre l'eterocromatina ha un aspetto più compatto ed è trascrizionalmente inattiva.
Questi termini furono proposti per la prima volta nel 1928 dal botanico tedesco Emil Heitz, in riferimento alla diversa intensità di colorazione che la cromatina di cellule di muschio assumeva durante il ciclo cellulare. Egli indicò come eterocromatina la parte che nel nucleo si colorava intensamente per tutta l'interfase, eucromatina quella che si colorava più debolmente.
Negli anni successivi si scoprì che eucromatina ed eterocromatina differivano funzionalmente e strutturalmente.
L'eucromatina rappresenta circa il 90% di tutta la cromatina e rispetto ad essa l'eterocromatina occupa posizioni differenti nel nucleo, soprattutto lungo la periferia.
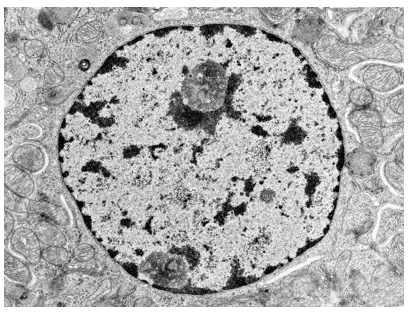
Immagine al microscopio elettronico a trasmissione che mostra nei dettagli le aree occupate e la diversa intensità di colorazione dell'eucromatina e dell'eterocromatina. L'eucromatina occupa quasi il 90% del nucleo ed avendo una struttura meno compatta assume una colorazione più chiara rispetto all'eterocromatina, che è variamente distribuita oltre che lungo la periferia.
Differenze strutturali tra eucromatina ed eterocromatina
L'eucromatina coincide con la porzione di cromatina le cui fibre hanno uno spessore di 30 nm.
L'eterocromatina presenta un grado di compattazione superiore a quello della fibra da 30 nm, in larga parte ancora da definire. È probabile che un numero maggiore di proteine intervenga in questo livello di organizzazione, contribuendo a rendere l'eterocromatina inaccessibile a tutti i fattori coinvolti nell'espressione genica.
L'eterocromatina si distingue in costitutiva e facoltativa. La prima rimane tale in tutte le cellule e in tutte le fasi del ciclo cellulare e si trova, ad esempio, nelle regioni del centromero e dei telomeri, di cui ne garantisce l'integrità strutturale. La seconda interessa quelle porzioni di cromatina in cui sono presenti geni che non devono essere espressi, a seconda del tipo di cellula, della fase del ciclo cellulare e del tipo di attività metabolica in corso.
Per la sua organizzazione strutturale, l'eterocromatina svolge anche una funzione protettiva dall'inserimento di elementi genetici mobili.
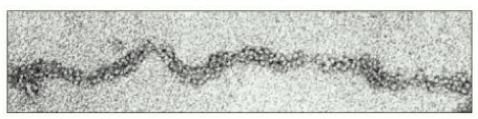
Immagine al microscopio elettronico che mostra la fibra da 30 nm della cromatina, isolata da nuclei interfasici. Essa rappresenta l'organizzazione strutturale dell'eucromatina. L'eterocromatina si presenta in uno stato più compatto, ancora non pienamente definito, in cui è coinvolto un numero maggiore di proteine.
Elementi fondamentali nella struttura della cromatina
La compattazione del DNA è resa possibile dall'intervento di numerose proteine, distinte in istoni e proteine non istoniche.
Gli istoni sono proteine basiche, cariche positivamente per l'alto contenuto di aminoacidi come lisina e arginina. Sono in grado di neutralizzare le cariche negative del DNA e consentire l'avvicinamento di più porzioni di esso senza che si respingano.
Intervengono sin dal primo livello di compattazione della cromatina, quello in cui essa appare come una collana di perle. Questa struttura può essere osservata al microscopio elettronico se la fibra da 30 nm in cui si presenta la cromatina estratta dai nuclei interfasici viene trattata con appositi agenti che ne causano l'apertura.
Nella struttura a collana di perle, il filo è rappresentato dal DNA, le perle dai nucleosomi.
Ogni nucleosoma ha un diametro di 10 nm ed è un complesso di otto istoni attorno al quale si avvolge un tratto di DNA lungo 146 pb (paia di basi). Gli istoni presenti sono indicati come H2A, H2B, H3 e H4, e di ognuno di essi sono presenti due copie. Il DNA che lega un nucleosoma all'altro è detto DNA linker o DNA nudo e ha una lunghezza variabile da poche paia di basi a circa 80.
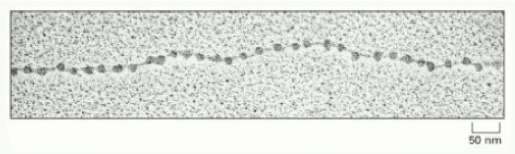
Immagine al microscopio elettronico che mostra la struttura a collana di perle del DNA, osservabile dopo trattamento della fibra da 30 nm con opportuni agenti lisanti. La collana di perle rappresenta la base dell'organizzazione strutturale della cromatina. Il filo è il DNA, le perle sono i nucleosomi.
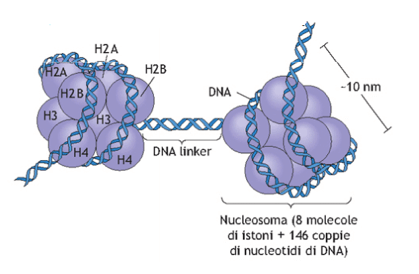
Ogni nucleosoma è un complesso di otto istoni (due copie per ogni istone H2A, H2B, H3 e H4) attorno al quale si avvolge un tratto di DNA lungo circa 146 paia di basi. Il DNA che unisce i vari nucleosomi è detto DNA linker.
Formazione della fibra da 30 nm dell'eucromatina
Come detto, se si estrae la cromatina dai nuclei di cellule in interfase, essa si presenta al microscopio elettronico formata da fibre dallo spessore di 30 nm. Se trattata in modo da ridurne il grado di compattamento, la fibra da 30 nm si svolge nella tipica organizzazione base della collana di perle.
La fibra da 30 nm corrisponde all'organizzazione strutturale dell'eucromatina e per la sua formazione sono stati proposti diversi modelli.
Uno dei modelli più accreditati è quello detto a ZigZag, di cui esisterebbero diverse varianti in funzione della lunghezza del DNA linker. Secondo questo modello, uno dei primi step nella formazione della fibra da 30 nm coinvolgerebbe un'altra tipologia di istone, l'istone H1. Esso agirebbe come una pinza legandosi tanto al DNA quanto alla componente proteica di due nucleosomi adiacenti, avvicinandoli. Questo processo sarebbe favorito anche dalle code degli istoni di ciascun nucleosoma.
La struttura della fibra da 30 nm è dinamica, in quanto i nucleosomi possono muoversi lungo il DNA per rendere o meno libera una determinata porzione di DNA. Questo processo è reso possibile da opportune modifiche covalenti delle code N-terminali degli istoni e dall'interazione dei nucleosomi con complessi multiproteici di rimodellamento della cromatina.
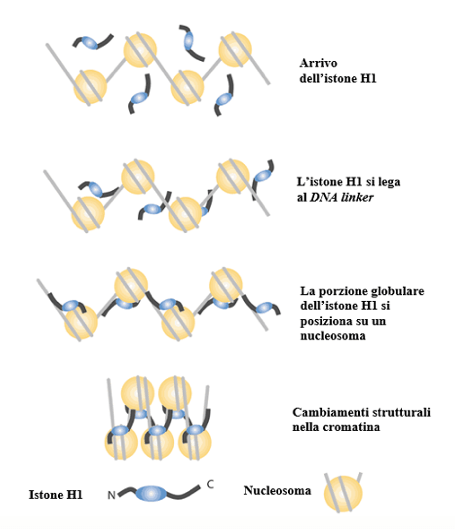
L'immagine mostra il ruolo dell'istone H1 nel promuovere la formazione delle fibre da 30 nm della cromatina.
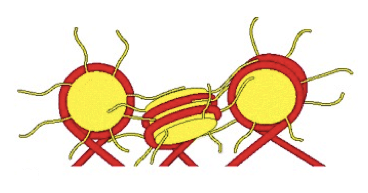
Nella formazione della fibra da 30 nm e quindi nel garantire l'avvicinamento dei nucleosomi e il reciproco contatto sarebbero coinvolte anche le code dei nucleosomi.
Possibili spiegazioni alla formazione dell'eterocromatina
L'eterocromatina ha un grado di compattazione maggiore rispetto all'eucromatina e rappresenta circa il 10% della cromatina del nucleo di una cellula eucariotica.
Il grado di compattamento dell'eterocromatina, ancora non ben definito, è superiore a quello della fibra da 30 nm e richiede l'intervento di numerose altre proteine.
Un modello proposto a seguito di studi condotti sui telomeri di cromosomi di lievito prevede il coinvolgimento di proteine chiamate Sir, in grado di interagire con i nucleosomi in cui le code N-terminali degli istoni H4 sono deacetilate. Anche la metilazione dei residui di lisina 9 dell'istone H3 avrebbe una funzione segnale nei confronti della formazione dell'eterocromatina. La funzione compattante delle proteine Sir sarebbe inoltre coadiuvata da specifiche proteine di legame al DNA.
Studi condotti sulla struttura dei centromeri dei cromosomi di lievito hanno invece indicato come cruciale nella formazione dell'eterocromatina una variante dell'istone H3.
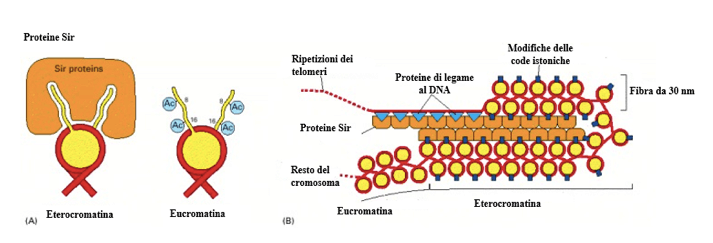
Modello per l'organizzazione strutturale dell'eterocromatina nei telomeri. In A la deacetilazione delle code istoniche favorirebbe il legame delle proteine Sir al nucleosoma. L'eucromatina si caratterizza, invece, per la presenza di residui acetilici sulle code istoniche. In B è possibile notare come anche il contributo di alcune specifiche proteine di legame al DNA (triangoli in azzurro) promuovi l'ulteriore compattazione della fibra da 30 nm cooperando con le proteine Sir.
Studia con noi