Cromosomi
Struttura e funzioni dei cromosomi
Per cromosoma si intende una struttura formata da DNA e proteine.
Un cromosoma veicola parte o tutto il patrimonio genetico di un organismo.
I cromosomi sono visibili soprattutto nelle cellule animali e nelle cellule vegetali quando vanno incontro a mitosi o meiosi e si presentano come dei corpiccioli simili a dei bastoncelli osservabili al microscopio ottico.
Sebbene il termine cromosoma sia specifico per le cellule eucariotiche, esso è usato anche per descrivere le strutture DNA-proteine delle cellule procariotiche.
I cromosomi procariotici hanno, tuttavia, un'organizzazione notevolmente diversa rispetto a quella dei cromosomi eucariotici.
I cromosomi delle cellule eucariotiche
Nelle cellule eucariotiche il genoma è ripartito in più cromosomi lineari, il cui numero varia notevolmente tra gli organismi senza che vi sia una correlazione con la complessità degli organismi stessi o con le dimensioni del loro genoma.
Ad esempio, il lievito ha 16 cromosomi, nel moscerino della frutta ne sono presenti 4.
I cromosomi sono visibili a partire dalla profase della mitosi e della meiosi nelle cellule coinvolte in questi processi.
Nelle cellule che non si dividono e durante l'interfase delle cellule che lo fanno, non sono visibili i cromosomi, ma una struttura meno compatta e organizzata in zolle e granuli che si chiama cromatina, anch'essa formata come i cromosomi da DNA e proteine.
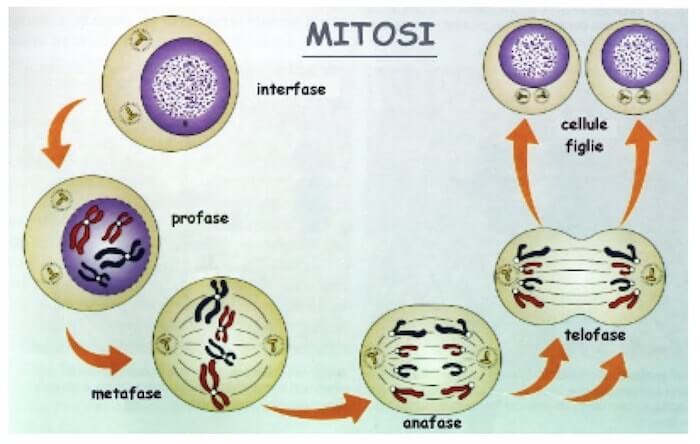
All'inizio della mitosi e della meiosi (profase) la cromatina osservabile in interfase assume un aspetto più compatto e si rendono visibili i cromosomi.
La cromatina
Il termine cromatina deriva dal greco "chroma", colore, per via delle proprietà tintoriali nei confronti di alcuni coloranti basici che rendono visibile il nucleo cellulare nei preparati istologici osservati al microscopio ottico.
Le proteine più importanti sono gli istoni, presenti in rapporto 1:1 rispetto al DNA.
Gli istoni sono proteine basiche per via della presenza prevalentemente di aminoacidi basici, cioè carichi positivamente a pH fisiologico, come arginina e lisina.
L'intima associazione tra DNA e istoni è resa possibile dall'interazione di natura elettrostatica tra le cariche negative del DNA dovute ai gruppi fosfato e quelle positive degli istoni, che svolgono quindi un ruolo importante nell'architettura della cromatina e dei cromosomi nel nucleo.
Oltre agli istoni, sono presenti diverse proteine, definite non-istoniche e presenti in rapporto variabile rispetto al DNA, coinvolte nella replicazione e riparazione del DNA, nell'espressione genica e nella sua regolazione.
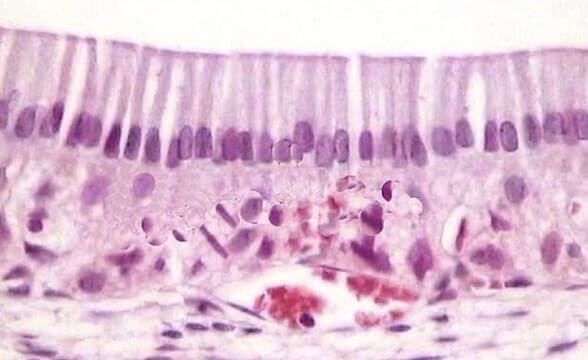
La cromatina deve il suo nome alla capacità di colorarsi intensamente. In questa immagine relativa a un preparato istologico di tessuto umano osservato al microscopio ottico sono chiaramente visibili i nuclei delle cellule.
Architettura della cromatina
Grazie agli istoni e ad alcune proteine non-istoniche il quantitativo totale di DNA (ogni cellula umana ne contiene circa 2 metri) può essere impacchettato nel nucleo che ha un diametro di soli 6 μm.
Gli istoni sono responsabili del primo livello dell'organizzazione strutturale dei cromosomi, rappresentato dal nucleosoma, scoperto nel 1974 in seguito ad esperimenti con l'enzima nucleasi.
Questo enzima agiva, degradandolo, solo su alcune regioni di DNA, lasciando supporre che la parte rimasta intatta fosse associata a proteine.
Quando il nucleo in interfase viene rotto delicatamente e il contenuto osservato al microscopio elettronico, la cromatina si presenta come un insieme di fibre di 30 nm di spessore.
In seguito al trattamenti con agenti che ne causano il rilassamento, la cromatina ha l'aspetto di una collana di perle.
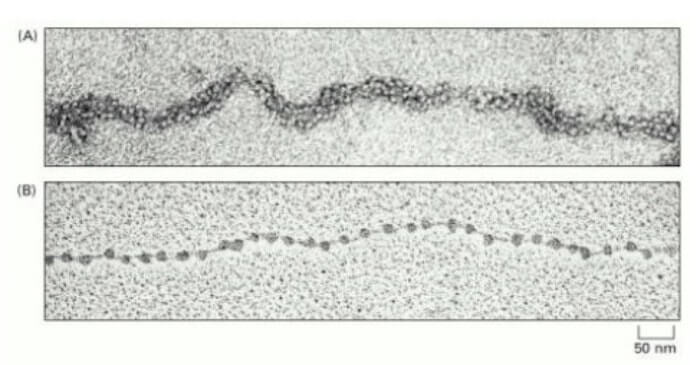
Nell'immagine A è visibile la fibra di 30 nm isolata dai nuclei interfasici, mentre in B si nota la struttura a collana di perle, in cui sono chiaramente visibili i nucleosomi e i tratti di DNA linker.
Le perle sono rappresentate dai nucleosomi, formati da un tratto di DNA lungo circa 146 bp (paia di basi) avvolto attorno a un core proteico contenente otto istoni, due per ogni istone H2A, H2B, H3 e H4.
Tra un nucleosoma e l'altro è presente un tratto di DNA nudo (non associato a proteine), detto DNA linker, la cui lunghezza media è di circa 50-70 bp.
L'unità ripetitiva della struttura a collana di perle, DNA avvolto e DNA linker, è pertanto di circa 200 bp, come era stato anche dedotto dagli esperimenti con la nucleasi.
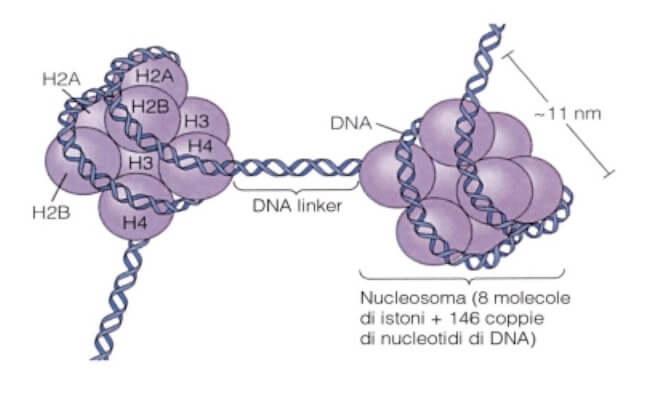
La struttura del nucleosoma con l'ottamero istonico e il tratto di DNA attorno ad esso.
Ruolo dell'istone H1
Oltre agli istoni dell'ottamero ne sono presenti altri, come l'istone H1, la cui funzione non è ancora del tutto nota.
Si ipotizza il coinvolgimento nel mantenimento dell'interazione tra DNA e istoni nel nucleosoma, oppure di avvicinamento di un nucleosoma all'altro per realizzare la struttura più compatta della fibra di 30 nm, che si ritiene sia la forma più comune presente nei nuclei interfasici.
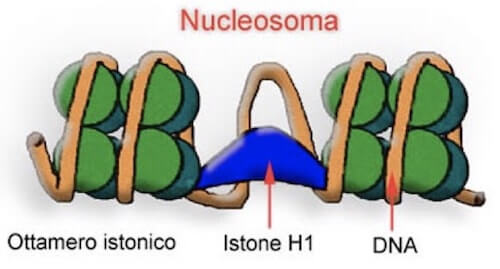
Possibile ruolo dell'istone H1 nella riduzione della distanza tra nucleosomi adiacenti nella fibra di 30 nm.
Formazione dei cromosomi
Quando avviene la divisione del nucleo in mitosi o meiosi, la cromatina raggiunge il massimo grado di compattamento e si rendono visibili i cromosomi.
La formazione di questi corpiccioli assicura un'equa ripartizione del materiale genetico tra le cellule figlie, difficilmente realizzabile in caso di un'organizzazione più lassa e meno compatta.
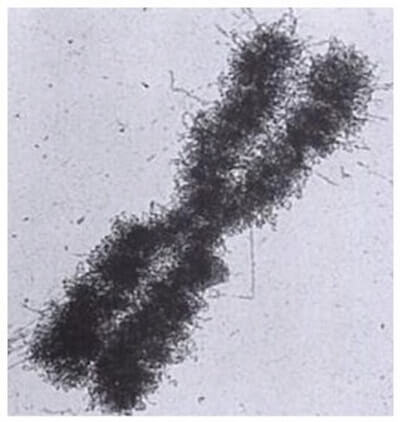
Cromosoma umano al microscopio elettronico.
Si ipotizza che l'aumento del grado di compattamento della cromatina si realizzi a partire dalla formazione sulla fibra di 30 nm di anse di 40-90 kb (chilobasi) di DNA, bloccate alla base da una struttura proteica indicata come scaffold nucleare.
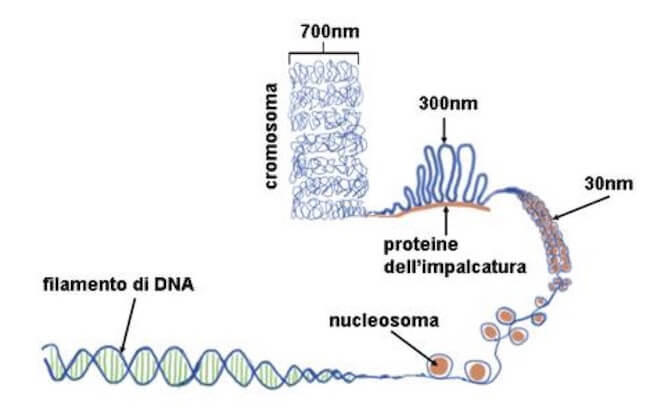
Stadi di compattamento sempre maggiori della cromatina. Lo spessore di 700 nm rappresenta il massimo grado di compattamento presente nei cromosomi.
Gli elementi distintivi dei cromosomi
Poiché le divisioni nucleari avvengono dopo la replicazione del DNA (che si realizza durante la fase S del ciclo cellulare), ciascun cromosoma si presenta raddoppiato.
Le due copie sono dette cromatidi fratelli, tenute insieme in corrispondenza del centromero, la cui posizione è diversa tra i vari cromosomi.
I cromosomi possono essere quindi riconosciuti in base alla posizione del centromero e alla lunghezza dei cromatidi.
I cromosomi in cui il centromero è centrale sono detti metacentrici; se il centromero è eccentrico e i bracci dei cromatidi sono ineguali, il cromosoma è submetacentrico o subtelocentrico, a seconda della maggiore o minore distanza rispetto alla parte terminale del cromosoma; con il centromero sull'estremità i cromosomi sono acrocentrici o telocentrici.
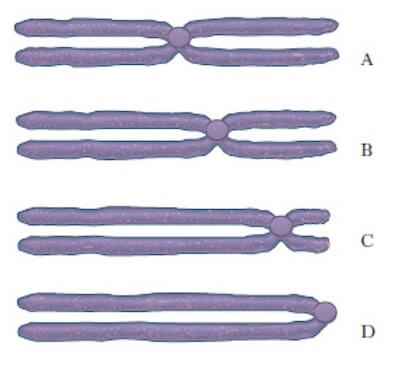
Vari tipi di cromosomi in base alla posizione del centromero. A) Cromosoma metacentico; B) Cromosoma submentacentrico; C) Cromosoma subtelocentrico; D) Cromosoma acrocentrico
In corrispondenza del centromero è presente un complesso proteico, il cinetocore, a cui si agganciano le fibre del fuso mitotico che assicurano il corretto movimento dei cromosomi durante la mitosi e la meiosi.
Un'altra importante regione dei cromosomi è quella terminale.
Le estremità dei cromosomi sono chiamate telomeri, dal greco telos, fine, costituite da DNA ripetitivo (sono presenti sequenze nucleotidiche molto lunghe ricche in TG).
I telomeri svolgono un'azione protettiva nei confronti di enzimi che potrebbero intaccare il DNA, assicurano la corretta replicazione dell'estremità dei cromosomi e collaborano con il sistema di riparazione del DNA.
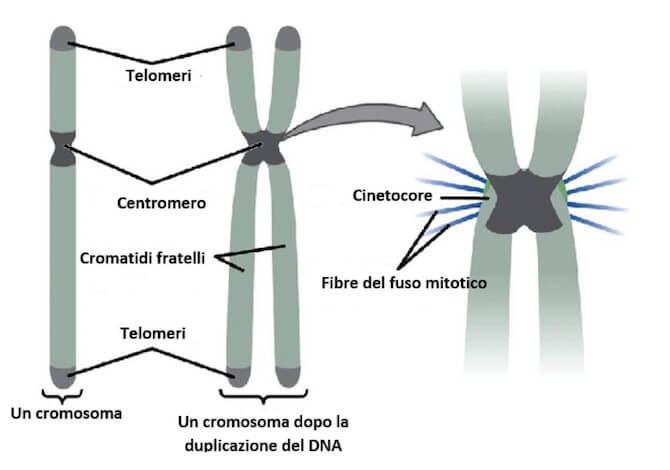
Gli elementi distintivi di un cromosoma.
Il numero di cromosomi è tipico di ogni specie
Il numero e la morfologia dei cromosomi è tipico di ogni specie. Nella maggior parte degli organismi eucariotici l'assetto cromosomico è diploide (condizione indicata con la sigla 2n), sono cioè presenti due copie dello stesso cromosoma. In molte piante il corredo è invece poliploide, con più copie dello stesso cromosoma.
I cromosomi omologhi
Nelle specie diploidi le due copie di ciascun cromosoma sono dette cromosomi omologhi, pertanto si parla di coppie di omologhi.
Nelle cellule dell'uomo sono presenti 46 cromosomi o 23 coppie di cromosomi omologhi.
Omologo significa "avere la stessa successione di geni", per cui ogni gene è presente in un organismo diploide in duplice copia.
Le due versioni di un dato gene in una coppia di cromosomi omologhi non devono essere necessariamente uguali e quindi omologo non significa uguale.
Il corredo cromosomico è organizzato in coppie di cromosomi perché al momento della formazione di un nuovo organismo, nelle specie che si riproducono per via sessuata, metà dei cromosomi sono forniti dal gamete maschile (spermatozoo), metà da quello femminile (cellula uovo).
A questo scopo i gameti sono aploidi (condizione indicata con la sigla n), contengono cioè metà dei cromosomi rispetto alle cellule somatiche. Il dimezzamento del patrimonio genetico necessario alla formazione dei gameti si realizza con la meiosi.
I cromosomi sessuali
Nell'uomo e negli altri organismi in cui il sesso è determinato geneticamente, le cellule dei due sessi differiscono per la morfologia di una coppia di cromosomi, detti sessuali, mentre gli altri cromosomi, detti autosomi, sono uguali nei due sessi.
Nei Mammiferi, in alcuni Anfibi e Pesci e in vari Invertebrati, gli individui di sesso femminile presentano due cromosomi sessuali uguali, detti XX, mentre nei maschi i due cromosomi sessuali sono diversi, uno X (si veda: cromosoma X) e uno piccolo Y.
Nei maschi, quindi, le coppie di omologhi sono 22, la coppia 23 è costituita da eterocromosomi.
Negli Uccelli, nei serpenti e in alcuni Invertebrati, sono le femmine a presentare cromosomi sessuali diversi, Z e W, mentre i maschi hanno cromosomi sessuali ZZ.
Il cariotipo
Lo studio del corredo cromosomico si realizza in massima parte durante la metafase della mitosi, quando i cromosomi sono meglio identificabili al microscopio ottico.
A questo scopo le cellule sono trattate con sostanze alcaloidi, come colchicina o vinblastina, che distruggono le fibre del fuso mitotico e bloccano la cellula in metafase.
I cromosomi vengono quindi colorati con vari metodi e le piastre metafasiche vengono fotografate. Le immagini ottenute sono utilizzate per creare il cosiddetto cariotipo: l'insieme dei cromosomi di una cellula, allineati in ordine di lunghezza decrescente e in base alla forma.
Grazie all'osservazione del cariotipo è possibile analizzare eventuali alterazioni nel numero o nella struttura dei cromosomi.
Lo studio del cariotipo e delle eventuali anomalie cromosomiche è proprio della citogenetica.
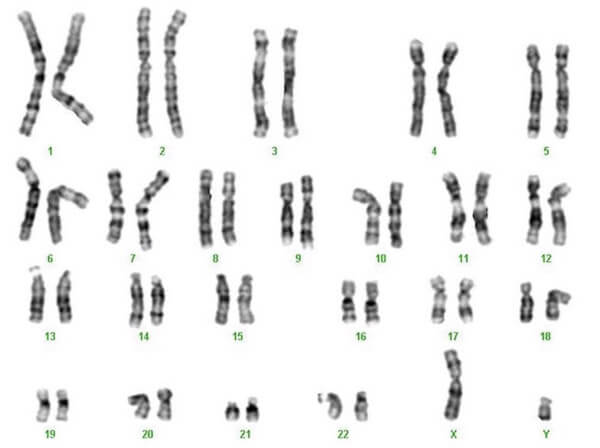
Cariotipo umano maschile. L'ultima coppia di cromosomi è rappresentata dai cromosomi sessuali X e Y. Nella donna sono presenti due cromosomi X.
I cromosomi procariotici
Il genoma di un procariote tipico, come Escherichia coli e molti dei batteri più studiati, è organizzato in una singola molecola circolare di DNA localizzata all'interno del nucleoide, l'unica regione che si colora leggermente in una cellula procariotica.
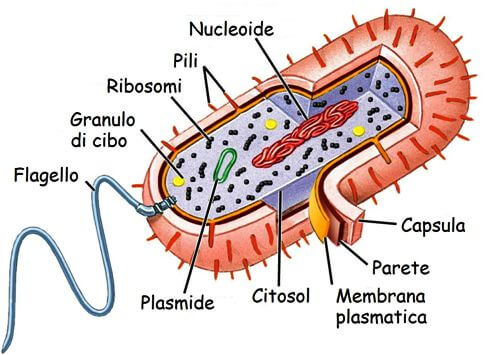
Ricostruzione di una generica cellula procariotica, con in evidenza la regione del nucleoide.
L'organizzazione del genoma procariotico in un'unica molecola circolare di DNA non è l'unico modello riscontrabile nei procarioti.
Sono sempre più numerosi gli esempi di genomi organizzati in una o più molecole lineari. Il primo cromosoma lineare è stato scoperto nel 1989 in Borrelia burgdoferi, il batterio che causa la malattia di Lyme, e, successivamente, scoperte simili sono state fatte in Streptomyces coelicor, un batterio del suolo, e in Agrobacterium tumefaciens, responsabile di un tipo di tumore, la galla del colletto, in alcune piante.
Il cromosoma di Escherichia coli
Il cromosoma di Escherichia coli ha una circonferenza di 1,6 mm e, come per i cromosomi eucariotici, deve essere compattato per rientrare nella cellula la cui circonferenza è di appena 2 μm.
Il compattamento viene raggiunto grazie all'associazione con proteine e ad avvolgimenti addizionali nella molecola DNA che la rendono superavvolta. Il superavvolgimento è reso possibile dall'intervento di due enzimi, la DNA girasi e la topoisomerasi I.
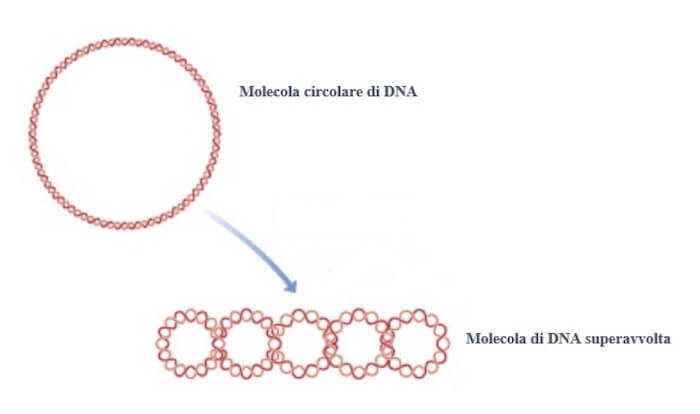
Il cromosoma circolare di Escherichia coli subisce una prima forma di impacchettamento mediante l'introduzione di superavvolgimenti.
Il modello attuale dell'organizzazione del cromosoma di Escherichia coli prevede un core proteico da cui si dipartono 40-50 anse superavvolte, ognuna delle quali contiene circa 100 kb di DNA.
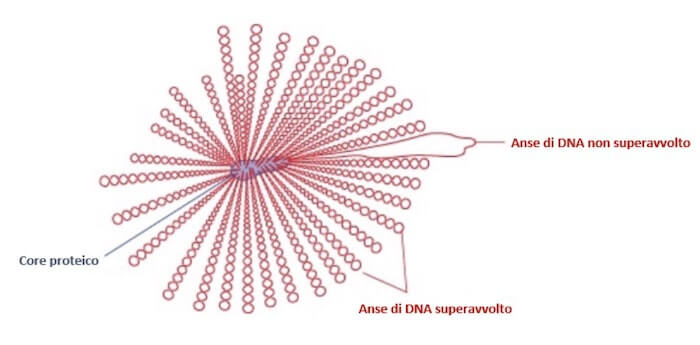
Un possibile modello dell'organizzazione del nucleoide in Escherichia coli. Circa 40-50 anse di DNA superavvolto si dipartono da un core proteico. Anche un singolo taglio può disavvolgere un'intera ansa.
La parte proteica del nucleoide contiene oltre agli enzimi DNA girasi e topoisomerasi I, anche proteine coinvolte nell'impacchettamento e con una funzione simile a quella degli istoni delle cellule eucariotiche.
La più abbondante di queste proteine è HU di cui sono presenti circa 60000 copie organizzate in tetrameri attorno a cui si avvolgono 60 bp di DNA.
La distribuzione e il distanziamento dei tetrameri non sono ancora noti.
Il cromosoma degli archeobatteri
Negli archeobatteri l'impacchettamento del DNA è reso possibile dall'intervento di proteine simili agli istoni che formano dei tetrameri, la cui associazione a circa 80 bp di DNA forma strutture simili ai nucleosomi dei cromosomi eucariotici.
Link correlati:
Quanti sono i cromosomi che ha l'uomo?
Quali possono essere gli effetti fenotipici delle mutazioni geniche?
Che cosa si intende per dosaggio genico?
Che cosa sono i trasposoni?
Che cosa sono gli enzimi isomerasi?
Che cosa sono le mutazioni cromosomiche?
Studia con noi